| Major Groups > Boletes > Suillus > Suillus cothurnatus |

|
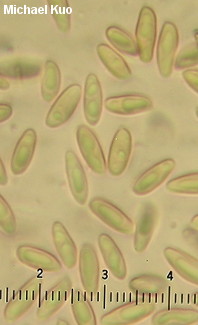
[ Basidiomycota > Boletales > Suillaceae > Suillus . . . ] Suillus cothurnatus by Michael Kuo, 17 November 2022 The morphological differences between Suillus cothurnatus and Suillus salmonicolor are few and far between—and, frankly, pretty unreliable. Both species have orangish-brownish, slimy caps and sheathing, glutinous rings that often feature a flaring, white, lower edge when the mushrooms are young. Some authors claim that the white, baggy edge of the ring is more prominent in Suillus salmonicolor, but I have not found this to be the case. Differences in size, how orange the flesh is, and in chemical reactions, are also put forth as definitive, but in my experience these do not appear to correlate with each other, let alone with species. There is also a putative micro-morphological difference (involving cystidia on the stem surface) recorded by some authors, but I have doubts about this, too. DNA study by Nguyen and collaborators (2016) suggests the possibility that Suillus cothurnatus and Suillus salmonicolor are the same, since specimens identified as both species formed a group together but did not separate into two clearly defined sub-groups. However, the authors were unable to study the type collections and definitive support for the synonymy was therefore elusive. Ultimately the two species (if indeed there are two, rather than one) might best be separated on the basis of their mycorrhizal partners, in my humble opinion: Suillus salmonicolor associates with jack pine, Virginia pine, and pitch pine, while Suillus cothurnatus associates with loblolly pine and longleaf pine (possibly also slash pine). Description: Ecology: Mycorrhizal with loblolly pine and longleaf pine (possibly also slash pine); growing alone, scattered, or gregariously; late summer and fall, or over winter in warmer climates; originally described from Florida (Singer 1945); widely distributed where the host trees occur naturally (from Mexico through Texas to Florida and New Jersey) and in plantations outside of this range; also reported from the Caribbean, Central America, and South America—as well as South Africa and Australia. The illustrated and described collections are from Illinois and Georgia. Cap: 2–6 cm; convex at first, becoming broadly convex; slimy when fresh, but often dry and glossy at maturity; bald, or very finely appressed-fibrillose and appearing streaked under the gluten; dull brownish orange to orangish yellow, with brownish streaks, becoming brownish to yellowish with maturity; the margin at first inrolled. Pore Surface: At first covered with a thick, whitish to orangish partial veil that is baggy and rubbery, with a white roll of tissue on the lower edge; orangish at first, maturing to yellowish or brownish; not bruising; 1–3 angular pores per mm; not boletinoid; tubes to about 1 cm deep. Stem: 3–6 cm long; 0.5–1 cm thick; tapered to base; tough; covered with glandular dots that are brownish red at first but become brown to black with age; whitish to yellowish or orangish; with a thick, sheathing, gelatinous, whitish to orangish ring that often features a whitish roll of tissue at the bottom and, in age, collapses to form a grayish, bracelet-like band; basal mycelium whitish to pastel orange. Flesh: Yellowish to orangish or orange in cap; darker orange in the stem; deep salmon orange in the stem base; often mottled; not staining on exposure. Odor and Taste: Not distinctive. Chemical Reactions: Ammonia negative to faintly purplish on cap surface; pinkish to purplish on flesh. KOH purple to gray on cap surface; purple to bluish on flesh. Iron salts negative on cap surface; negative to faintly greenish on flesh. Spore Print: Cinnamon brown. Microscopic Features: Spores 7–10 x 2–3 µm; boletoid-fusiform; smooth; yellowish in KOH. Basidia 22–28 x 4–6 µm; clavate; 4-sterigmate. Cystidia in gelatinized bundles, often poorly defined individually; 35–50 x 5–8; cylindric to subclavate; thin-walled; smooth; dark brown in KOH. Pileipellis an ixolattice of poorly defined elements 2.5–5 µm wide, smooth, hyaline in KOH. REFERENCES: R. Singer, 1945. (Smith, Smith & Weber, 1981; Palm & Stewart, 1986; Both, 1993; Bessette, Roody & Bessette, 2000; Klofac, 2013; Bessette et al., 2016; Nguyen et al., 2016; Woehrel & Light, 2017.) Herb. Kuo 07300908, 10170902, 10141301. This site contains no information about the edibility or toxicity of mushrooms. |
© MushroomExpert.Com |






|
Cite this page as: Kuo, M. (2022, November). Suillus cothurnatus. Retrieved from the MushroomExpert.Com Web site: http://www.mushroomexpert.com/suillus_cothurnatus.html |