Microscopic Details
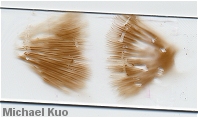
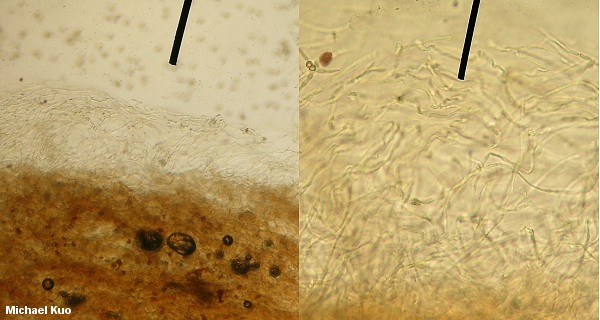
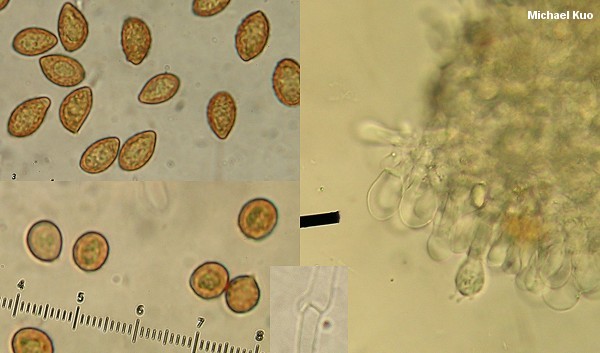
Microscopic examination is generally required for identification of all but a few "field guide species" in the genus. Spore morphology is probably the most crucial among the microfeatures; spore shape, size, and the degree of ornamentation (from nearly smooth to strongly verrucose) can all be important in identification. Ideally, spores should be measured from a spore print so that they are unquestionably mature. A Roman aqueduct section, mounted in 2% KOH, will suffice for studying other microscopic features important in Cortinarius, which include the pileipellis (see the discussion and illustration above, under "Sliminess"), the presence or absence of clamp connections, and cystidia on the gills. Impressive, well defined cystidia are almost never heard of in Cortinarius (Cortinarius violaceus is virtually the only exception), but some species possess cylindrical to club-shaped cheilocystidia--sometimes designated as "marginal cells" when they are so inconspicuous as to be unworthy of a full-blown "cheilocystidia" designation (yes, the distinction is frustrating, and no, authors do not use these terms consistently and universally).
Species Pages
Cortinarius alboviolaceus
Cortinarius anomalus
Cortinarius armillatus
Cortinarius azureus
Cortinarius bolaris
Cortinarius caesiocanescens
Cortinarius caperatus
Cortinarius collinitus
Cortinarius corrugatus
Cortinarius croceus
Cortinarius distans
Cortinarius elegantio-montanus
Cortinarius glaucopus
Cortinarius hesleri
Cortinarius infractus
Cortinarius iodeoides
Cortinarius iodes
Cortinarius malicorius
Cortinarius marylandensis
Cortinarius mucosus
Cortinarius multiformis
Cortinarius olearioides
Cortinarius ophiopus
Cortinarius pinguis
Cortinarius privignoides
Cortinarius rubripes
Cortinarius sanguineus
Cortinarius seidliae
Cortinarius semisanguineus
Cortinarius smithii
Cortinarius species 01
Cortinarius species 02
Cortinarius subpulchrifolius
Cortinarius torvus
Cortinarius trivialis
Cortinarius vibratilis
Cortinarius violaceus
References
Ammirati, J. F. & Smith, A. H. (1969). Studies in the genus Cortinarius, I: Section Dermocybe, Cortinarius aureifolius complex. The Michigan Botanist 8: 175–180.
Ammirati, J. F. (1972). The section Dermocybe of Cortinarius. Ph. D. thesis, University of Michigan. 282 pp.
Ammirati, J. F. & Smith, A. H. (1972). Studies in the genus Cortinarius, II: Section Dermocybe, new and interesting species from Michigan. The Michigan Botanist 11: 13–25.
Ammirati, J. F. (1975). Cortinarius, section Dermocybe-�Cortinarius clelandii. Mycotaxon 3: 95–101.
Ammirati, J. F. & Smith, A. H. (1977). Studies in the genus Cortinarius, III: Section Dermocybe, new North American species. Mycotaxon 5: 381–397.
Ammirati, J. F. & A. H. Smith (1978). Studies in the genus Cortinarius, IV: Section Dermocybe, new North American species. Mycotaxon 7: 256–264.
Ammirati, J. F. & H. E. Bigelow (1984). Cortinarius iodes versus Cortinarius heliotropicus. Mycotaxon 20: 461–471.
Ammirati, J. F. & Smith, A. H. (1984). Cortinarius II: A preliminary treatment of species in the subgenus Dermocybe, section Sanguinei, in North America, north of Mexico. McIlvainea 6: 54–64.
Ammirati, J. F. (1988). Dermocybe (or Cortinarius, subgenus Dermocybe): Some species in the sections Dermocybe, Holoxanthae, and Malicoriae. McIlvainea 8: 48–59.
Ammirati, J. F. (1988). Dermocybe (Cortinariaceae, Agaricales): North American names - new combinations, synonyms, and notes. Mycotaxon 33: 437–446.
Ammirati, J. F. (1989). Dermocybe, subgenus Dermocybe, section Sanguineae in northern California. Mycotaxon 34: 21–36.
Ammirati, J. F., K. W. Hughes, K. Liimatainen, T. Niskanen & P. B. Matheny (2012). Cortinarius hesleri from eastern North America and related species from Europe and western North America. Botany 91: 91–98.
Brandrud, T. E., H. Lindström, H. Marklund, J. Melot & S. Muskos (1990). Cortinarius: Flora Photographica. Volume 1. Matfors, Sweden: Cortinarius HB.
Brandrud, T. E., H. Lindström, H. Marklund, J. Melot & S. Muskos (1992). Cortinarius: Flora Photographica. Volume 2. Matfors, Sweden: Cortinarius HB.
Brandrud, T. E., H. Lindström, H. Marklund, J. Melot & S. Muskos (1994). Cortinarius: Flora Photographica. Volume 3. Matfors, Sweden: Cortinarius HB.
Breitenbach, J. & F. Kränzlin (2000). Fungi of Switzerland: A contribution to the knowledge of the fungal flora of Switzerland. Volume 5. Agarics Part 3. Cortinariaceae. Lucerne: Edition Mykologia. 338 p.
Cripps, C. L. & Miller, O. K. Jr. (1994). A new Cortinarius from a mature aspen stand in Montana. Mycotaxon 50: 315–321.
Froslev, T. G., T. S. Jeppesen & T. Laessoe. (2006). Seven new calochroid and fulvoid species of Cortinarius. Mycological Research 110: 1046–1058.
Garnica, S., M. Weiss, B. Oertel & F. Oberwinkler (2003). Phylogenetic relationships of European Phlegmacium species (Cortinarius, Agaricales). Mycologia 95: 1155–1170.
Garnica, S., P. Spahn, B. Oertel, J. Ammirati & F. Oberwinkler (2011). Tracking the evolutionary history of Cortinarius species in section Calochroi, with transoceanic disjunct distributions. Evolutionary Biology 11: 213.
Hansen, L. & Knudsen, H., eds. (1992). Nordic macromycetes Vol. 2: Polyporales, Boletales, Agaricales, Russulales. Copenhagen: Nordsvamp. 474 pp.
Hoiland, K. & Holst-Jensen, A. (2000). Cortinarius phylogeny and possible taxonomic implications of ITS rDNA sequences. Mycologia 92: 694–710.
Judge, B. S., J. F. Ammirati, G. H. Lincoff, J. H. Trestrail III & P. B. Matheny (2010). Ingestion of a newly described North American mushroom species from Michigan resulting in chronic renal failure: Cortinarius orellanosus. Clinical Toxicology 48: 545–549.
Kauffman, C. H. (1906). Cortinarius as a mycorhiza-producing fungus. Botanical Gazette 42: 208–214.
Kauffman, C.H. (1918). The gilled mushrooms (Agaricaceae) of Michigan and the Great Lakes region, Volumes I and II. New York: Dover. 924 pp. (1971 Reprint.)
Kauffman, C. H. (1932). Cortinarius. North American Flora 10: 282–348.
Knudsen, H. & J. Vesterholt, eds. (2008). Funga Nordica: Agaricoid, boletoid and cyphelloid genera. Copenhagen: Nordsvamp. 965 pp.
Liu, Y., S. O. Rogers, J. F. Ammirati & G. Keller (1995). Dermocybe, section Sanguineae: A look at species relationships within the sanguinea complex. Sydowia 10: 142–154.
Liu, Y. J., S. O. Rogers & J. F. Ammirati (1997). Phylogenetic relationships in Dermocybe and related Cortinarius taxa based on nuclear ribosomal DNA internal transcribed spacers. Canadian Journal of Botany 75: 519–532.
Moser, M. (1983). Keys to Agarics and Boleti (Polyporales, Boletales, Agaricales, Russulales). Ed. Kibby, G. Transl. Plant, S. London: Roger Phillips. 535 pp.
Moser, M. (1993). Studies on North American Cortinarii. III. The Cortinarius flora of dwarf and shrubby Salix associations in the alpine zone of the Windriver Mountains, Wyoming, USA. Sydowia 45: 275–306.
Moser, M. M., K. H. McKnight & J. F. Ammirati (1995). Studies on North American Cortinarii I. New and interesting taxa from the greater Yellowstone area. Mycotaxon 55: 301–346.
Moser, M. M. & Ammirati, J. F. (1996). Studies in North American Cortinarii II. Interesting and new species collected in the North Cascade Mountains, Washington. Mycotaxon 58: 387–412.
Moser, M. M. & J. F. Ammirati (1997). Studies on North American Cortinarii IV. New and interesting Cortinarius species (subgenus Phlegmacium) from oak forests in northern California. Sydowia 49: 25–48.
Moser, M. M. & J. F. Ammirati (1999). Studies in North American Cortinarii V. New and interesting Phlegmacia from Wyoming and the Pacific Northwest. Mycotaxon 72: 289–321.
Moser, M. M. (2002). Studies in the North American Cortinarii VII. New and interesting species of Cortinarius subgen. Telamonia (Agaricales, Basidiomycotina) from the Rocky Mountains. Feddes Repertorium 113: 48–62.
Niskanen, T. (2008). Cortinarius subgenus Telamonia in North Europe. Doctoral dissertation, University of Helsinki. Helsinki, Finland. Available online: http://urn.fi/URN:ISBN:978-952-10-4446-5.
Niskanen, T., I. Kytovuori & K. Liimatainen (2011). Cortinarius sect. Armillati in northern Europe. Mycologia 103: 1080–1101.
Niskanen, T., I. Kytövuori, E. Bendiksen, K. Bendiksen, T. E. Brandbrud, T. G. Frøslev, K. Høiland, T. S. Jeppesen, K. Liimatainen & H. Lindström (2018). Cortinarius (Pers.) Gray. In Knudsen, H. & J. Vesterholt, eds. (2018). Funga Nordica: Agaricoid, boletoid, clavarioid, cyphelloid and gastroid genera. Copenhagen: Nordsvamp. 762–885.
Peintner, U., E. Horak, M. Moser & R. Vilgalys (2002). Phylogeny of Rozites, Cuphocybe and Rapacea inferred from ITS and LSU rDNA sequences. Mycologia 94: 620–629.
Peintner, U., M. Moser & R. Vilgalys (2002). Thaxterogaster is a taxonomic synonym of Cortinarius: New names and new combinations. Mycotaxon 81: 177–184.
Peintner, U., M. Moser & R. Vilgalys (2002). Rozites, Cuphocybe and Rapacea are taxonomic synonyms of Cortinarius: New combinations and new names. Mycotaxon 83: 447–451.
Peintner, U., Moncalvo, J. M., & Vilgalys, R. (2004). Toward a better understanding of the infrageneric relationships in Cortinarius (Agaricales, Basidiomycota). Mycologia 96: 1042–1058.
Phillips, R. (1986). A provisional multi-access key to American species of Cortinarius, Inoloma & Telemonia (including Leprocybe, Sericeocybe and Hydrocybe). Published by the author. 45 pp.
Seidl, M. T. (2000). Phylogenetic relationships within Cortinarius subgenus Myxacium, sections Defibulati and Myxacium. Mycologia 92: 1091–1102.
Smith, A. H. (1939). Studies in the genus Cortinarius I. Contributions from the University of Michigan Herbarium 2. 43 pp.
Smith, A. H. (1942). New and unusual Cortinarii from Michigan, with a key to the North American species of subgenus Bulbopodium. Bulletin of the Torrey Botanical Club 69: 44–64.
Smith, A. H. & L. R. Hesler (1943). New and interesting agarics from Tennessee and North Carolina. Lloydia 6: 248–266.
Smith, A. H. (1944). New and interesting Cortinarii from North America. Lloydia 7: 163–235.
Smith, A. H. & Trappe, J. M. (1972). The higher fungi of Oregon's Cascade Head Experimental Forest and vicinity - I. The genus Phaeocollybia (Agaricales) and notes and descriptions of other species in the Agaricales. Mycologia 64: 1138–1153.
Soop, K. (1993). On Cortinarius in boreal pine forests. Agarica 12: 101–116.
This site contains no information about the edibility or toxicity of mushrooms.
Cite this page as:
Kuo, M. (2011, December). The genus Cortinarius. Retrieved from the MushroomExpert.Com Web site: http://www.mushroomexpert.com/cortinarius.html