| Major Groups > Boletes > Red-Pored Boletes > Boletus subvelutipes |

|
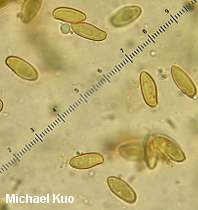
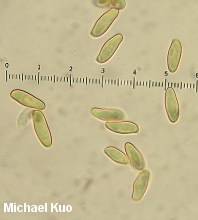
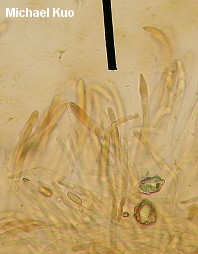
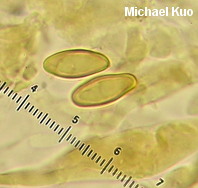
Boletus subvelutipes species group [ Basidiomycota > Boletales > Boletaceae > Boletus . . . ] by Michael Kuo I'm going to make this easy. If your brown-capped, red-pored, blue-staining bolete was collected under hardwoods east of the Rocky Mountains and has a stem that lacks reticulation but features a finely fuzzy coating over its base, call it "Boletus subvelutipes"--unless you are in the southeastern United States, in which case you should consider Boletus austrinus (stem covered with purple floccules) and Boletus hypocarycinus (basal mycelium white, spores under 12 µ long). That's pretty much as accurate as you can be--even from a scientific standpoint, unless by "scientific" you really mean, "I want to immerse myself in a mess centered around a 40-year-old treatise of Michigan boletes," or, "I want to apply European species names and concepts to North American boletes despite the fact that the European species probably don't actually occur on our continent." If any of that sounds like fun to you, I've done my best to help you out, starting with couplet #50 of my key to red-pored boletes. Alternatively, you could try entertaining yourself with Smith & Thiers's 1971 key to Stirps subvelutipes. (Alter-alternatively, you could just pound your head into a wall.) This is an area of bolete taxonomy that needs a lot of work. I encourage you to participate! If you find subvelutipes-like boletes in the woods, why not take careful notes on the ecology, photograph or scan the mushrooms, describe the collection, and preserve your specimens? Described below, and illustrated to the right, are examples representing three rather different subvelutipes-like boletes I have collected. Description of 09070701: Ecology: In grass at woods' edge, with white oak, northern red oak, slippery elm, shagbark hickory, and sassafras nearby; growing gregariously; September 7, 2007; Coles County, Illinois, USA. Cap: 6-11 cm; convex, becoming broadly convex to flat; dry or sticky; bald; medium brown (OAC 735), fading somewhat (OAC 729); dark brown after drying. Pore Surface: When young pinkish orange (OAC 693) with yellow areas; when mature dingy orange-brown; bruising promptly blue; depressed at stem, even in buttons; pores angular, 2 per mm at maturity; tubes to 15 mm deep. Stem: 45-60 x 9-12 mm; slender; equal; tough; bright yellow under dense red floccules; becoming brownish basally; bruising blue; base velvety, crimson. Flesh: Yellow in cap; bright yellow in stem; staining quickly blue on exposure. Odor and Taste: Not distinctive. Chemical Reactions: Ammonia negative to orangish on cap; erasing blue to orange on flesh. KOH red on cap; orange on flesh. Iron salts grayish on cap and flesh. Spore Print: Olive. Microscopic Features: Spores 12-15 x 4.5-5.5 µ; fusiform; yellowish in KOH; smooth. Hymenial cystidia infrequent; fusiform; not projecting. Pileipellis a trichoderm of densely packed cylindric elements 2.5-5 µ wide; terminal elements cylindric with rounded apices; hyaline to orangish brown or brownish orange in KOH. Caulocystidia fusiform; hyaline to brownish in KOH; 35-50 µ long. Description of 06190401 and 06221002: Ecology: Under oaks, maples, and hickories; growing gregariously; June; Richmond and Jasper counties, Illinois, USA. Cap: 2-10 cm; convex, becoming broadly convex; dry; velvety; when fresh olive (OAC 783, 784), with orangish, yellow, or red areas; soon more or less brown to olive brown; bruising strongly and quickly blue to blackish; margin initially inrolled; dark brown after drying. Pore Surface: Bright red with a yellow marginal zone at first, becoming reddish orange; eventually yellowish orange and finally dingy olive; bruising quickly dark indigo to nearly black; 3-4 pores per mm; tubes to 1 cm deep. Stem: 50-60 x 15-20 mm; equal, with a slightly rooting tapered base; not reticulate, or becoming slightly reticulate near the apex; yellow at the apex; red below; discoloring brownish with age; bruising quickly blue to black; base velvety, yellow to red. Flesh: Yellow in cap; dark yellow in stem; sometimes red in stem base; staining quickly blue on exposure. Odor and Taste: Not distinctive. Chemical Reactions: Ammonia negative to reddish on cap; erasing blue to yellow on flesh. KOH mahogany red on cap; orange on flesh. Iron salts blackish to dark grayish green on cap; negative to grayish on flesh. Spore Print: Olive. Microscopic Features: Spores 10-12 x 3.5-4.5 µ; fusiform; yellowish in KOH; smooth. Hymenial cystidia inconspicuous; fusoid-ventricose; hyaline to yellowish in KOH; to 40 µ long. Pileipellis a trichoderm of cylindric elements 2.5-5 µ wide; subterminal elements yellowish in KOH; terminal elements golden (creating a contrasting upper turf), cylindric with subacute apices; all elements brown in Melzer's. Caulocystidia fusoid-ventricose to subfusiform; yellow to golden in KOH; to 40 µ long. Note: In many ways these collections match what used to be called "Boletus erythropus" and is now known as Boletus luridiformis. These, however, are European species. Description of 06140802: Ecology: Solitary under white oak and sycamore; June 14, 2008; Madison County, North Carolina, USA. Cap: 5 cm; convex; dry; suedelike; yellow brown, with a yellow marginal zone; bruising blue; dingy yellow after drying. Pore Surface: Orangish red with a yellow marginal zone; bruising quickly blue; 3-4 pores per mm; tubes about 1 cm deep. Stem: 100 x 20 mm; tapering slightly to base; not reticulate; yellow at the apex; dull brownish below; bruising blue; base velvety, dark red. Flesh: Yellow; staining blue on exposure. Odor and Taste: Not distinctive. Chemical Reactions: Not recorded. Spore Print: Not obtained. Microscopic Features: Spores 11-17 x 4.5-6 µ; fusiform; yellow in KOH; smooth. Hymenial cystidia abundant; fusoid-ventricose; yellow in KOH; to 50 µ long. Pileipellis a cutis of cylindric elements 2.5-12 µ wide; hyaline to yellow in KOH. Caulocystidia fusoid-ventricose; yellow in KOH; 35-65 x 10-15 µ. Note: Of the collections described on this page, this one agrees most closely with Peck's Boletus subvelutipes, as interpreted by Smith & Thiers (1971): its microscopic features match well, as do the yellow brown colors and the yellow dried cap. REFERENCES: Peck, 1889. (NOTE: It does not appear to me that all of the sources listed here are describing the same species. Saccardo, 1891; Singer, 1945; Smith & Thiers, 1971; Grund & Harrison, 1976; Smith, Smith & Weber, 1981; Lincoff, 1992; Both, 1993; Horn, Kay & Abel, 1993; Barron, 1999; Bessette, Roody & Bessette, 2000; Roody, 2003; McNeil, 2006; Miller & Miller, 2006; Binion et al., 2008.) This site contains no information about the edibility or toxicity of mushrooms. |
|








|
Cite this page as: Kuo, M. (2013, December). Boletus subvelutipes species group. Retrieved from the MushroomExpert.Com Web site: http://www.mushroomexpert.com/boletus_subvelutipes.html ©MushroomExpert.Com |